Time weaves memories into patterns

When I get a craving for Cote d’Or chocolate, I always hit up London Drugs. It’s not the only grocer around that carries the brand, nor is the chocolate always in stock. But based on past experiences, I know I have a much better chance at finding the decadent treat there than anywhere else.
In a constantly changing world, how do we know what’s normal? Psychological and computer modeling studies suggest that our brains extract commonalities from similar experiences as we go about our daily lives. Individual memories are thus generalized and integrated into a knowledge scaffold – a pattern, if you will. When a new experience comes along, the brain parses out differences between existing and novel information, and updates the relevant memory scaffold to better reflect reality. Rather than relying on individual occurrences, we understand the world through patterns.
But what are the brain computations that weave individual memories into a big-picture perspective of the world? Does pattern recognition occur immediately after laying down a memory, or does it only happen with the passage of time?
Emerging patterns
Researchers trained mice to find a platform in a pool of water, and moved the platform daily following a specific rule. Although the platform seemed to bounce around randomly, a pattern emerged across multiple training days, in that the platform tended to appear more often in some locations than others.
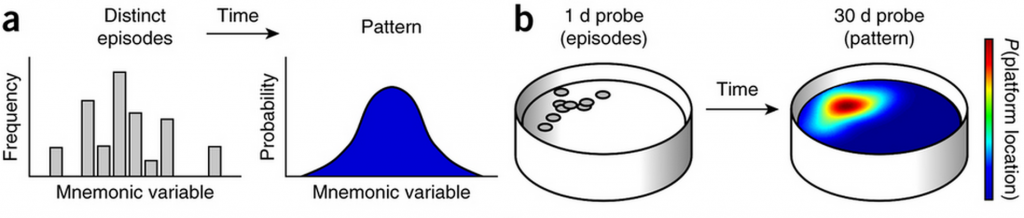
Figure 1. Distinct experiences morph into a statistical model. The warmer the colour, the higher the probability of the platform appearing.
After the last training session, researchers waited either 1 or 30 days before placing the mice back into the pool and watched them navigate to their old stomping grounds. With only a day’s break, the mice frequently swam across places where they had previously found the platform – in particular, the location of the last platform. After 30 days, however, the mice changed their search strategy: rather than zeroing in on any particular learned location, they focused their search to areas of the pool where the platform had most frequently appeared. Somehow, with the passage of time, the mice had extracted a pattern from their training day memories to predict where the platform would most likely be. Control experiments ruled out other factors that may have caused the observed shift in search strategy, such as decreased memory accuracy.
Researchers next tested whether the same results hold for a more difficult pattern. This time they moved the platform around in the north and east half of the pool, but was twice as likely to place it in the north than east quadrant (see below). One day after training, the mice spent a similar amount of time navigating both quadrants; after 30 days, however, they swam according to the pattern of platform locations. The mice spent roughly twice as much time in the north than east quadrant, and also lingered in places where the platform had a high chance of appearing.

Figure 2. Left: warmer colours indicate more time spent in the area. Right: 1 day after training, the mice spent a roughly equal amount of time in the North and East quadrant (N/E ratio nearly 1). 30 days later they spent twice as much time in the North than East quadrant (N/E ratio roughly 2).
Updating patterns
Once we identify patterns that reflect average (i.e “normal”) occurrences, anything that doesn’t fit immediately flags our attention, letting us know that our knowledge framework needs to be updated. To see if the passage of time increases the ability to identify outliers, researchers trained a new group of mice on the same platform-finding task as the first experiment. After the last training day, they waited 1 or 30 days and then introduced the mice to a new platform location, which either fitted the original pattern or appeared somewhere completely unexpected. Think rain in Vancouver (fits the pattern) versus rain in the Sahara desert (that’s just weird).

Figure 3. The newly introduced platform is either consistent (black) or conflicting (white) with the original pattern.
Interestingly, congruency mattered for both time delays: the mice had no trouble finding the location of the consistent platform, but struggled to find the unexpected one. So it seems that the brain is keeping tabs even during the initial learning, and that the mice may already have an inkling that a pattern existed. However, the difference was much more pronounced 30 days after the initial training. This suggests that once a pattern emerges from individual memories, it influences our choices and behaviours: we don’t go searching for rain in the Sahara because we don’t expect to find it there. In other words, we become more adept at picking out unusual occurrences that don’t fit.
When something farfetched happens it forces us to re-evaluate and update our model of the world. Thus an unexpected platform location should strongly promote learning in mice, but only after sufficient time had passed to allow the identification of a pattern. This was indeed the case: mice that were given a 30 day break after their initial training dramatically altered their search strategy to include the new information. They also learned the unusual platform location much better than those that only had a day off.
Learning outliers
How does the brain process new information that conflicts with established knowledge? Previous studies suggest that the medial prefrontal cortex (mPFC) may be involved. To test this idea, researchers expressed an off-switch for neuronal activity specifically in the mPFC of mice. The switch is a specialized receptor that only responds to a synthetic chemical; injecting the chemical silences neurons. This technique, called inhibitory DREADDs, allows researchers to rapidly turn off neuronal activity whenever they please.
The researchers waited 30 days after training to challenge the mice with a new platform location that conflicted with the original pattern. In one group of mice, they silenced mPFC neuronal activity before the challenge (labelled "CNO" in the graph below). When tested a day later, control mice changed their search strategy to incorporate the new and surprising information – they repeatedly circled the new position (top right graph in d). But this behaviour was lost in mice with inhibited mPFC function: they spent less time at the new platform location, instead returning to areas they had frequented during training (bottom right graph in d), seemingly loosing any sensitivity to the unexpected. In stark contrast, mPFC inhibition had no effect on learning if the new platform position fitted the original pattern, and did not impact learning if the mice were only allowed a day’s break after initial training.

Figure 6. In d, left column is consistent platforms (black), right is conflicting ones (white). Warmer colours represent more time spent in the area. f: after inhibiting the mPFC with CNO, the mice spent less time near the new conflicting platform (white bar on the far right). CNO did not affect learning for consistent platforms (left two graphs, compare white to black bar).
In sum, the brain seems to recognize statistically common elements across distinct memories. With the passage of time, these elements are woven into an overall understanding of the world. With these models, we can predict the probability of different events based on past experiences. When our predictions fail, the mPFC helps to resolve the conflict by incorporating new information into our models. And thus we are able to keep up with the constantly changing world.
Richards BA, Xia F, Santoro A, Husse J, Woodin MA, Josselyn SA, & Frankland PW (2014). Patterns across multiple memories are identified over time. Nature neuroscience PMID: 24880213






